The 'virus' or the egg?
Which came first?


A common objection raised by the defenders of virology (see for example Steve Kirsch) in response to those who say that ‘viruses’ ‘don’t exist’ is that ‘viruses’ exist, because we have photographs of them. For example, the following photograph shows particles seen under an electron microscope that scientists have called ‘West Nile virus’.
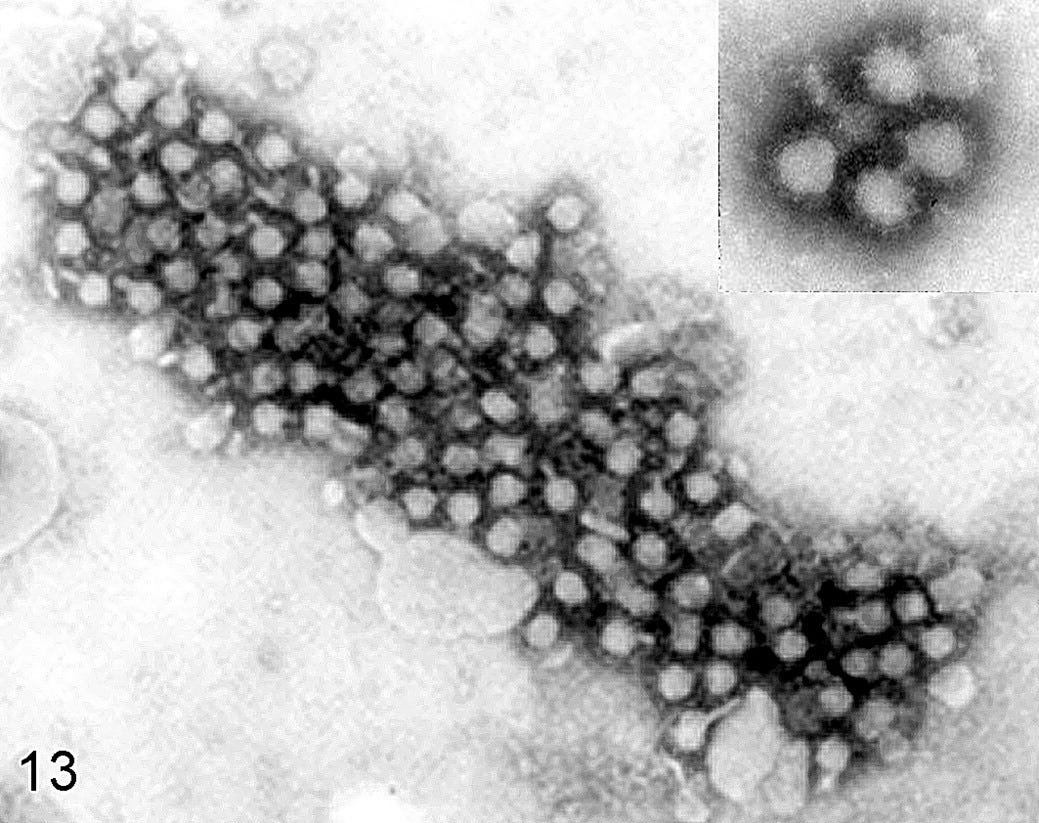
There are thousands of photographs like this that can be found for any given ‘virus’, as well as thousands of colourful, computer-generated graphics of these particles. The following image is a rendering of the aforementioned ‘West Nile virus’.

It goes without saying that a picture of a unicorn, convincing as it may be, does not automatically prove that unicorns are real, physical entities.

A photo of ‘Nessie’ does not prove that a prehistoric sea creature lives at the bottom of Loch Ness.

On the other hand, we know that berries exist because, not only do we have photographs of them, we can also pick them off bushes or buy them from our local grocery store. However, a photograph of a berry does not tell us whether that particular type of berry is poisonous.
: Black nightshade vs deadly nightshade")
In short; photographs of particles that are labelled ‘virus’ do not automatically mean that the particles in question have the characteristics that are attributed to them, namely, the ability to cause ‘disease’. Thus, additional steps need to be taken to demonstrate that they do indeed, possess such characteristics. We previously covered these in more detail in ‘What is Germ Theory?’.
It is worth noting at this stage that when scientists take photographs of cells and other materials they are viewing under their electron microscopes, they are already causing important structural changes. An article published in 2016, titled ‘Live Cell Electron Microscopy Is Probably Impossible’ states that “the minimal needed electron dose to obtain contrast is already many orders of magnitude above the lethal dose known to cause reproductive-cell death”. Additionally, various cytotoxic fixing and staining agents are used in an attempt to preserve the appearance of the cells before viewing them under a microscope. According to an article published on electron microscope manufacturer Leica’s website, “the most widely used stains in electron microscopy are the heavy metals uranium and lead”.
How do these methods impact what scientists are viewing under their microscopes? The work of Harold Hillman (1930 – 2016), a Reader of Physiology and director of the Unity Laboratory of Applied Neurobiology at the University of Surrey is particularly interesting in this regard. Hillman, like many others before and after him, fell from grace when in the early 1960s, he “found that when centrifuged, adenosine triphosphate [ATP] changed its activity. This meant that many experiments probing this crucial substance could be wrong” (The Scientist, 1988).
Hillman couldn’t get his results published, and as appears to be standard procedure, he was attacked and smeared by the medical orthodoxy and its followers, whom Hillman adequately likened to “a religion, with its priesthood and theology”. Perturbed by his findings and their reception by his colleagues, Hillman began to investigate other experimental techniques. In 1972, he published ‘Certainty and Uncertainty in Biochemical Techniques’. According to The Scientist “the reviews were almost uniformly hostile. The most common complaint was that the arguments were old hat—the problems of preparation artifects and degradation had all been solved or corrected for in the early days of the techniques. However, no one has answered to Hillman’s satisfaction the questions of when these confirmatory tests were ever done and where they were published. He believes to this day that no one has ever bothered to do the control experiments”. This apparent lack of proper controls appears to be a common problem in this field of study.
A couple of years later, whilst doing work on nerve cells, Hillman and microscopist Peter Sartory used transmission electron microscopy to examine the cells in question and “were astonished by what they saw”;
“We noticed something so peculiar that we really couldn’t’ believe it,” Hillman recalls. “About 80% of the membranes in the cell appeared end-on, [as if the cell had been sliced through its center]. It took Sartory and myself several weeks to realize that it simply wasn’t possible.” Yet that is what virtually all electron micrographs and illustrations in papers and textbooks show. “That was our first shock,” says Hillman. That implied that every electron microscopist was cutting virtually everything in the cell perfectly at right angles.”
According to conventional thinking, cell membranes consist of two layers. In stained EM sections the two layers show as two parallel lines of dark stain, like railroad tracks. But, Hillman observes, however you cut a cell, the two lines are always the same distance apart in the EM image. He draws the analogy of a chef cutting an orange. If he slices it clean through the center, the sliced surface will show a thin rim of peel. If he cuts it with a glancing blow, so that only a small slice is taken off, the peel won’t be cut at right angles—it will show as a much “thicker” rim. Hillman reasons that because the rim is always the same width in EM sections, it cannot represent a thick two-layer membrane. Rather, he believes, it is a single thin membrane stained on both sides.
“I have challenged electron microscopists to make a three-dimensional model of any living cell in which this [a membrane appearing to have identical thickness however it is cut] is so. It simply isn’t possible. All the stains they examine them with are heavy metals that deposit on both sides of the membrane, thus any real membrane will appear as two lines.”
According to an article published by The Telegraph, Hillman and Sartory “concluded that the processes of extracting, dehydrating and staining cells so that they can be seen under an electron microscope results in fundamental changes to the cells themselves. Thus, many of the subcellular structures observable under an electron microscope could be “artefacts” – a spurious result produced by the testing procedure, and not a biological reality”.
Others appear to recognise these problems. According to Audrey Glauert, head of electron microscopy at the Strangeways Research Laboratory in Cambridge; “every electron image is an artifact. With EM you’re a long way from the original cell – you’ve treated it and sectioned it and fixed it and done all kinds of things to it”. Glauert however, believes that Hillman and Sartory’s work is ‘misguided’, and “is particularly upset that in their quest to get published, Hillman and Sartory wrote their ideas up for School Science Review, a journal for science teachers”.
The ‘virus’ or the egg?
As we discussed in ‘A primer on virology and epidemiology’, a ‘virus’, according to Britannica, is “an infectious agent of small size and simple composition that can multiply only in living cells of animals, plants, or bacteria. The name is from a Latin word meaning ‘slimy liquid’ or ‘poison’.”
According to an article published by National Geographic, “an estimated 10 nonillion (10 to the 31st power) individual viruses exist on our planet”. Interestingly, many appear to be part of us. According to an article published by Cold Spring Harbor Laboratory titled ‘The non-human living in you’; “8% of our DNA consists of remnants of ancient viruses, and another 40% is made up of repetitive strings of genetic letters that is also thought to have a viral origin”.
According to a paper published in Nature in 2021, “the human body also hosts vast numbers of different viruses, collectively termed the ‘virome’.” This so-called ‘virome’ consists of “approximately 10 [to the 13th power] particles per human individual, with great heterogeneity”. The paper further states that “in recent years, studies of the human virome using metagenomic sequencing and other methods have clarified aspects of human virome diversity at different body sites, the relationships to disease states and mechanisms of establishment of the human virome during early life. Despite increasing focus, it remains the case that the majority of sequence data in a typical virome study remain unidentified, highlighting the extent of unexplored viral ‘dark matter’”.
These differences that appear to relate to “different body sites” are interesting, given that different ‘viruses’ are said to “attack certain cells in your body such as your liver, respiratory system, or blood”. And perhaps this is where so-called ‘dormant viruses’ come from. An example of such a ‘virus’ is the ‘Varicella Zoster [VZ] virus’, which we previously discussed in some detail in ‘Chickenpox, shingles, smallpox, monkeypox and friends’. VZ is said to cause both ‘chickenpox’, and ‘shingles’. The virus is said to typically cause ‘chickenpox’ in young people, and then remains ‘dormant’ in the body. Later in life, it may become ‘re-activated’, and cause ‘shingles’. How ‘it’ is able to ‘survive’ in an apparently hostile environment is unclear, as are the reasons for its ‘re-activation’. According to the Mayo Clinic; “the reason for shingles is unclear. It may be due to lowered immunity to infections as people get older. Shingles is more common in older adults and in people who have weakened immune systems”. This in turn suggests that the ‘virus’ is not the primary problem, as we previously discussed here.
When ‘viruses’ replicate, they are said to cause cell death, which scientists use in cell culture experiments as indirect evidence of their disease-causing capabilities; “they invade living, normal cells and use those cells to multiply and produce other viruses like themselves. This can kill, damage, or change the cells and make you sick”.
‘Viruses’ cannot replicate outside of cells. According to an article published in National Geographic; “they rely on cellular machinery to help with their replication, so they need to have some sort of primitive cell to make use of that machinery," said Jack Szostak, a biochemist at Harvard University and a Nobel laureate.” In other words, viruses mooch off cells, so without cells, viruses can't exist”.
Individual ‘viral’ particles – ‘virions’ – are made up of a DNA or RNA ‘core’, enveloped by a ‘shell’ – called a ‘capsid’, itself made of protein. According to some, this ‘capsid’ is an important characteristic that differentiates ‘viruses’ from other similar particles. The following computer-generated graphic is a rendition of the icosahedral ‘capsid’ of an ‘adenovirus’. It is unclear how scientists are able to assert that the imperfect particles they are viewing under their microscopes look anything like this.


Given that ‘viruses’ need cells to replicate, this raises the question; what came first – the cell or the ‘virus’? According to an article in National Geographic, “the predominant theories for the origin of viruses propose that they emerged either from a type of degenerate cell that had lost the ability to replicate on its own or from genes that had escaped their cellular confines.”
It would appear therefore that the first ‘viruses’ did not ‘hijack’ the cell machinery of the cell, but rather, were created by the cell itself.
Extracellular vesicles
In an article published in 2003, the author notes that in regards to HIV “the virus is fully an exosome in every sense of the word”. ‘Exosomes’ are a subtype of ‘extracellular vesicles’ [EVs]. According to a paper published in 2019, “extracellular vesicles (EVs) are lipid bound vesicles secreted by cells into the extracellular space. The three main subtypes of EVs are microvesicles (MVs), exosomes, and apoptotic bodies, which are differentiated based upon their biogenesis, release pathways, size, content, and function”.
‘Exosomes’ appear to bear an uncanny resemblance to ‘viruses’. To illustrate these similarities, here are cryo-electron microscopy photos of ‘extracellular vesicles’, followed by photos of ‘coronaviruses’. It is difficult to argue that visually, they are not indistinguishable.


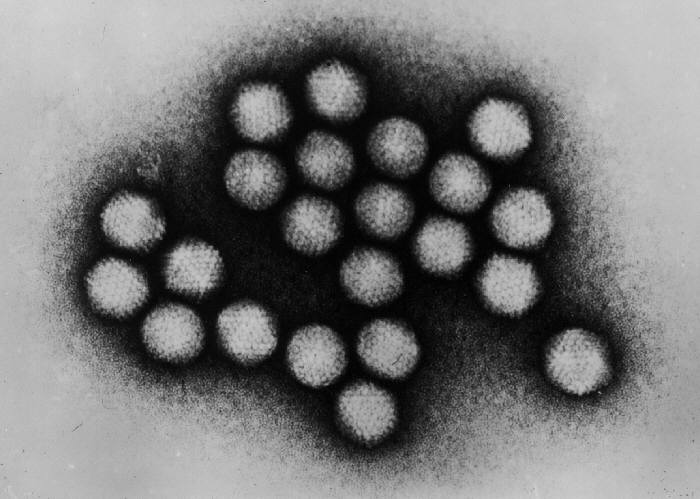{kind=link}
According to an article published by Cell Guidance Systems, they appear to share a great deal more in common; “exosomes and viruses have many things in common, including their size, lipid coat, and the ability to transport RNA and proteins between cells”. The so-called ‘capsid’ also appears to be present in ‘exosomes’. According to an article published in Quanta Magazine; “vesicles carry various RNA and proteins, including some that resemble viral proteins and can form capsid structures”.

Ultracentrifugation is a method that is commonly used to allegedly separate ‘virions’ (individual ‘viral’ particles) from other particles. However, according to Cell Guidance Systems, this technique cannot reliably distinguish ‘viruses’ from ‘exosomes’; “because exosomes and viruses are similar in size and density, separation of the two is not possible using this technique [ultracentrifugation]”. A paper published in the journal Viruses in 2020 confirms this; “nowadays, it is an almost impossible mission to separate EVs [extracellular vesicles] and viruses ... because they are frequently co-pelleted due to their similar dimension”.
We previously discussed the ‘cytopathic effect’, which is one of the key markers scientists look for when attributing the role of ‘virus’ to arbitrarily selected particles viewed under their electron microscopes. In a paper published in 2019, researchers used ‘exosomes’ that were extracted from cells that had been infected with the ‘Newcastle Disease virus’ to infect cells in cell culture. They observed a cytopathic effect; “to our surprise, the cytopathic effect was significantly observed in cells treated with exosomes from NDV[Newcastle disease Virus]-infected cells”.
It would appear therefore, that ‘exosomes’ are also able to cause a ‘cytopathic effect’, further blurring an already, very blurred line. ‘Exosomes’ also appear to contain ‘viral’ RNA. In a paper published in 2021, researchers found that “analysis of exosome content purified from critical and non-critical patients revealed the presence of SARS-CoV-2 RNA in the exosomal cargo”. They also appear to play an important role in the infection of cells. A paper published in 2019 found that “rabies virus infection increased the release of exosomes ... the role of exosomes in virus infection process remains unclear and needs to be further investigated”.
According to Cell Guidance Systems, the only real fundamental difference between ‘exosomes’ and ‘viruses’, is how they are produced: “by definition, a virus can replicate by using host machinery. Exosomes do not replicate in recipient cells. Each cell has an innate ability to make its own exosomes. Thus, an exosome or other EV cannot be a virus because they cannot replicate”.
Particles that are said to be created by ‘hijacking cell machinery’, are still created by the cell, as are those that aren’t created through this ‘hijacking’ mechanism. It is unclear therefore, how scientists are able to reliably tell the two apart. As previously discussed, electron microscopy kills the cells, and therefore this process has never, as far as we are aware, been observed in real-time, meaning this alleged difference appears to be purely theoretical. The ability to ‘hijack cells’ does not appear to be unique to ‘viruses’ either, as discussed in ‘Lost in Translation’.
Are ‘viruses’ contaminated ‘exosomes’?
In 2020, an article published in Nature on the subject of ‘bacterial exosomes’ noted that among other things, they may be involved in the transfer of toxins. Toxins such as arsenic can take the form of nanoparticles. This image shows arsenic trioxide particles in cell culture. They are very small, less than 10nm. In comparison, the Sars-COV-2 virus, has apparently been found to measure 50 – 140 nm.

This raises the question of whether nanoparticles such as these may be contained within the materials scientists are working with. According to the Nature article, it seems possible that some ‘exosomes’ may either contain toxins, which they may release along with the rest of their cargo, when ‘docking’ with other cells, or that certain toxins can bind to their surface proteins. Either of these would result in these ‘exosomes’ inadvertently spreading toxins throughout the body, in much the same way ‘viruses’ are said to work.
As we’re previously discussed in various articles, sulphur and phosphorus, among others, are essential elements found throughout the body. Arsenic readily binds to both, resulting in wide-ranging problems.
In ‘The Spectrum of Diabetes’, we found that arsenic has a clear diabetogenic effect as a result of its ability to bind with disulphide bridges found inside both insulin and insulin receptors, rendering them ‘non-functional’. In ‘Malaria or mal’aria?’, we hypothesised that this affinity may also explain how, for instance, mosquitoes and other insects may be able to contribute to the transfer of toxins, and therefore, spread of ‘disease’. Mosquito saliva has been found to contain more than a hundred proteins. Some of these, such as the anti-coagulant AAPP contain a number of sulphur-rich cysteine-residues. This in turn suggests that mosquitoes exposed to arsenic, when injecting their saliva during a blood meal, may also be injecting some of the arsenic they have bioaccumulated.
‘Viral load’ or a marker for toxicity?
In a study published in 2022, researchers in China found that; “Circulating exosome levels may be novel biomarker for ecotoxic effects of water environment”. In other words, they found higher levels of ‘exosomes’ in fish exposed to various pollutants, including arsenic and mercury.
Other studies appear to show similar findings. In a paper published in 2018, researchers found that there was a negative correlation between lung function in the case of ‘progressive fibrosing interstitial lung disease of unknown aetiology’ and levels of ‘exosomes’, leading the researchers to conclude that they are “promising biomarkers for diagnosis and disease severity”.
As mentioned previously, most ‘viral infections’ are asymptomatic, suggesting that the particles in question are not the fundamental problem. We have previously found a similar pattern both for bacteria and helminths such as plasmodium, said to be the cause of ‘malaria’. It follows therefore, that ‘viruses’ – or ‘exosomes’ may fall into the same category.
All of this may explain what the mysterious particles were that scientists discovered at the time of the ‘SMON’ epidemic which we have discussed in detail here. An apparently novel ‘disease’ who’s defining manifestation was an epidemic in Japan that began at the end of the 1950s, and lasted until approximately 1970, a number of microorganisms were implicated, including a novel ‘virus’. Eventually, it was established that the cause of the ‘disease’ was not the novel ‘virus’ – or any ‘virus’ or microorganism for that matter – but a drug, clioquinol. It is clear from reading the literature on the subject that clioquinol was not the fundamental cause of the problem. We will discuss this in more detail in a future article.
What is interesting about this case, is that the novel ‘virus’ that was discovered, known as the ‘SMON virus’, met all the requirements to demonstrate pathogenicity, but it was decided that it played no role. Some years later, in 1984, this same ‘virus’ was renamed ‘Inoue-Melnick virus’, and it was found in patients in the US suffering from multiple sclerosis. In 1993, one of the many ‘mysterious diseases’ that seem to regularly appear out of nowhere, swept through Cuba. This same ‘virus’ was once again found in samples taken from sick patients.
A word on ‘viral genomes’ and testing
The question of ‘viral genomes’ is often brought up as further evidence that the particles scientists are viewing under their microscopes are distinct. Given that the these particles are never properly isolated and purified, it is unclear how exactly these genomes are sequenced, and what it is they actually represent.
According to the NIH, PCR is a laboratory technique that “uses selective primers to 'copy' specific segments of a DNA sequence", which in turn are used to match "a segment of the virus's genetic material”. This suggests in turn that the tests are not specific and open to interpretation. According to PCR test manufacturer Labcorp with regards to their monkeypox assay: “although this assay does not differentiate vaccinia or monkeypox virus from cowpox, camelpox, ectomelia or gerbilpox virus, a positive result with this assay in the United States is most likely due to monkeypox virus or vaccinia virus; however, potential exposure to other Orthopoxviruses should be considered”.
This shows how testing protocols and the interpretation of the results can be adjusted to create the illusion that an alleged pathogen is no longer of concern, and has been ‘eradicated’ through ‘vaccination’ and other measures, which is what is said of ‘smallpox’ and to some extent, ‘polio’. As discussed in ‘Malaria or mal’aria?’, ‘malaria’ provides one of the best examples for how differences in testing protocols distract people with endless lists of diseases and alleged pathogens, and ignore the common denominator, which appears to be mass poisoning.
This doesn't mean the tests are not testing for something specific. As Caroline previously discussed in ‘Quick fire 1’, PCR testing can be used to test for certain markers such as γH2AX, which forms when double-strand DNA breaks appear. Exposure to ionising radiation is one cause, and therefore γH2AX is a marker for radiation poisoning. In ‘Unpicking the PCR’, we saw how an increase in the presence of γH2AX was observed in cells ‘infected’ with ‘Sars-COV-2’ (fluids taken from a sick person). An increase in other markers for DNA damage (ATR and CHK1) was also detected.
Given that it is the fluids or materials containing the ‘virus’ that are sequenced, rather than the particle itself, it seems possible and indeed, highly likely that when ‘viral’ genomes are sequenced, markers such as this are included in that sequence. This explains why a paper published in 2020 appears to have found ‘viral particles’ in samples taken from individuals who tested both positive and negative for ‘COVID’.

Different ‘viral’ genomes, composed from materials extracted from different parts of the body (‘the human virome’), may include markers for damage in different areas. In other words, these genomes are not really genomes at all – a genome typically maps back to a given entity. Rather, these are genomic sequences that can be thought of as a ‘snapshot’ of the state of materials extracted from a given organism that is out of homeostatic balance. The ‘imbalance’ is caused by exposure the aforementioned substances, which will in turn result in the presence of certain markers. As discussed in ‘A Hitchiker’s Guide to Exosomes’, it would appear that this is what the Sars-COV-2 genome represents.
Conclusion
It would appear that the particles scientists are calling ‘virus’ are in fact, in some cases, a marker for ‘disease’. As we have previously discussed, the very concept of ‘disease’ is questionable and could be better thought of as varying symptoms of chronic and acute poisoning that manifest in a wide range of seemingly unrelated conditions.
Given that most ‘viral infections’ are said to be ‘asymptotic’, and that certain ‘environmental factors’, namely, malnutrition and exposure to certain toxins are said to play an important role in determining whether a given infection becomes ‘symptomatic’, this would mean that all ‘viral diseases’ are in fact caused by some form or other of intoxication. The implications of this cannot be overestimated.
Thank you as always to Caroline for her invaluable help in pulling this information together. Thank you to Eva for sharing Hillman’s work, and Mike Stone for the many useful resources shared here.
 | A guest post by
|
This is a GREAT article. Clarified so much for me. Wish I could get others to read it and digest the facts it contains. Unfortunately, people seems to be comfortable with and cling to their delusions.
Thank you. A wonderful read.